Outline of Cardiovascular Morphogenesis
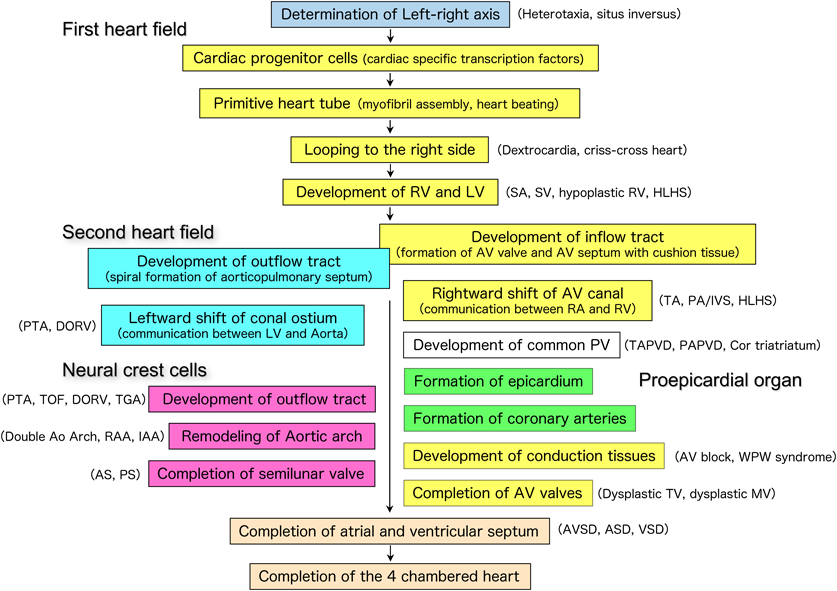
Embryonic heart development begins with the determination of the left-right axis in the primitive node. This information is transmitted to the left and right lateral plate mesoderm. Immature mesoderm cells differentiate into cardiac progenitor cells that express myocardial-specific transcription factors in the cardiac primordium (cardiogenic region). Myocardial progenitor cells gradually migrate to the middle of the embryo and differentiate into cardiac myocytes, forming a single primitive heart tube. The primitive heart tube starts to contract in a peristaltic motion, then gradually begins to rhythmically contract, forming the contour of the heart by bending to the right side of the embryo (cardiac looping). In the primitive heart tube, sinus venosus, primitive atrium, primitive ventricle, bulbus cordis, and truncus arteriosus, the segments are aligned from the caudal direction to the cranial direction. As the cardiac looping proceeds, these anteroposterior segments are converted into a left–right positional relationship, particularly in the ventricles. As the heart tube elongates, the vertical relationship of the atria and ventricles is exchanged. Inside the primitive heart tube, four endocardial cushions develop in the atrioventricular canal to form two atrioventricular valves and part of the atrioventricular septum. Two (four in the initial stages) conotruncal swellings develop in the outflow tract, dividing the pulmonary artery and aorta in a spiral manner. In humans, after the formation of the atrial and ventricular septum is completed on approximately embryonic day 50, the heart with two atria and two ventricles is finally completed1–5) (Fig. 1).
Cardiovascular morphogenesis is initiated by progenitor cells, including the first heart field cells, which mainly give rise to the cardiomyocytes of the left ventricle and part of the atria; the second heart field cells, which give rise to the cardiomyocytes of the right ventricle and part of the atria; cardiac neural crest cells which differentiate into the fibroblasts and smooth muscle cells of the aorta and pulmonary artery; and the proepicardial organ, which originates from the boundary with the liver and migrates into the looping heart tube to form the epicardium, which eventually gives rise to fibroblasts, and vascular endothelial and smooth muscle cells5) (Fig. 2).
Most congenital heart diseases result from either the suspension of or deviation from these processes of cardiovascular morphogenesis due to genetic abnormalities in the fetus and/or environmental factors in the fetus and mother.
Determination and Transmission of the Left-Right Axis Information: Heterotaxy (Asplenia, Polysplenia), Situs Inversus, and Kartagener Syndrome
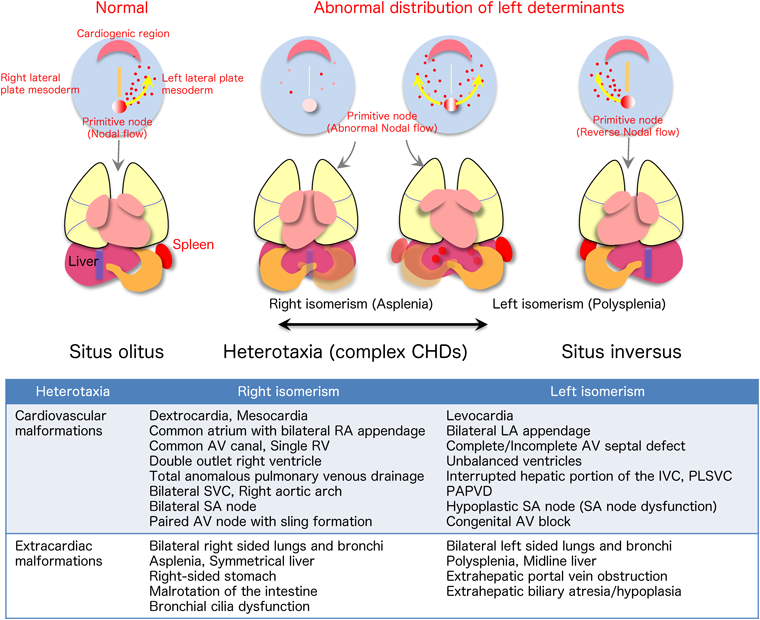
The node pit cells in the primitive node, located in the middle of the three-layered blastoderm, have cilia that rotate in a counterclockwise spiral. Because the cilia are inclined to align caudally, a leftward nodal flow occurs at the top of the node pit cells. Node crown cells sense this flow and upregulate the intracellular calcium level. Then a growth factor, Nodal, is predominantly expressed on the left side of the node, which in turn upregulates growth factors and transcription factors Nodal, Lefty, and Pitx2 in the left lateral plate mesoderm. Subsequently, signals are finally transmitted to the left side of the heart primordium (cardiogenic region). If the left/right determinants are not sufficient in the primitive node, or if the leftward nodal flow is not normal, the left/right information of the organs, including the heart, is not established or randomized. Heterotaxy, right isomerism (splenia) or left isomerism(polysplenia), may develop6) (Fig. 3).
Determination and Differentiation of Myocardial Cells
In the cardiogenic region located on the cranial portion of the embryo, immature mesodermal cells give rise to cardiac progenitor cells that only express cardiac specific transcription factors, such as Nkx2.5 and GAT A4. These cardiac progenitor cells are determined to become mature cardiomyocytes of the first (both the atrium and left ventricle) and second (both the atrium and right ventricle) heart fields. These progenitor cells form bilateral cardiac primordia and migrate to the middle of the embryo and eventually fuse to form a straight heart tube. At the same time, contraction proteins are expressed in the progenitor cells, which later differentiate into cardiac myocytes with mature myofibrils. Then, the primitive heart tube starts to contract rhythmically.1, 2)
Formation of the Heart Tube and Cardiac Looping: Atrioventricular Discordant, Congenitally Corrected Transposition of the Great Arteries, Dextrocardia, Superior-Inferior Ventricles, Crisscross Hearts, etc.
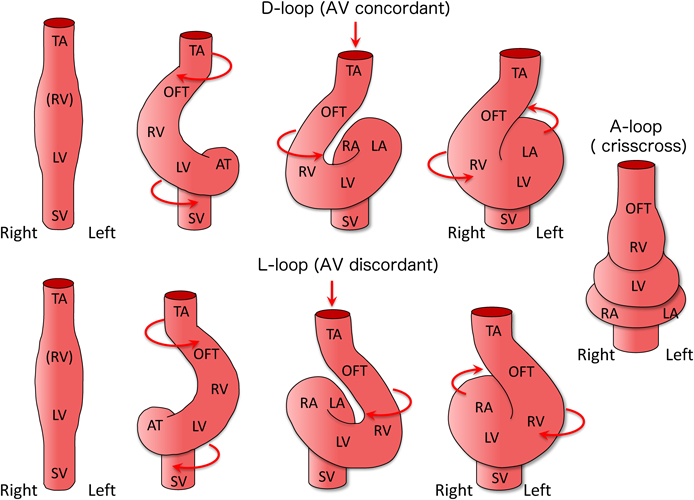
The primordia of the left and right ventricles rapidly bulge as the heart tube elongates. The straight heart tube starts to loop to the right side of the embryo, creating the outer contour of the four-chambered heart (Fig. 4). After completion of the looping process, the circulatory system of the embryo with a series circuit shifts to a parallel one, where the pulmonary and systemic circulations become separated and independent (Fig. 4). If the straight heart tube loops to the “left side” of the body, an inconsistent connection between the atria and ventricles takes place, resulting in “atrioventricular discordance”. Furthermore, when the heart tube loops in the front direction, the normal parallel atrioventricular connection changes to a vertical and torsional relationship, forming “superior-inferior ventricles”. If the atrioventricular connection is extremely twisted, “crisscross heart” is formed.5)
Rightward Shift of the Atrioventricular Canal: Tricuspid Atresia, Single Inlet of Univentricular Atrioventricular Connection, etc.
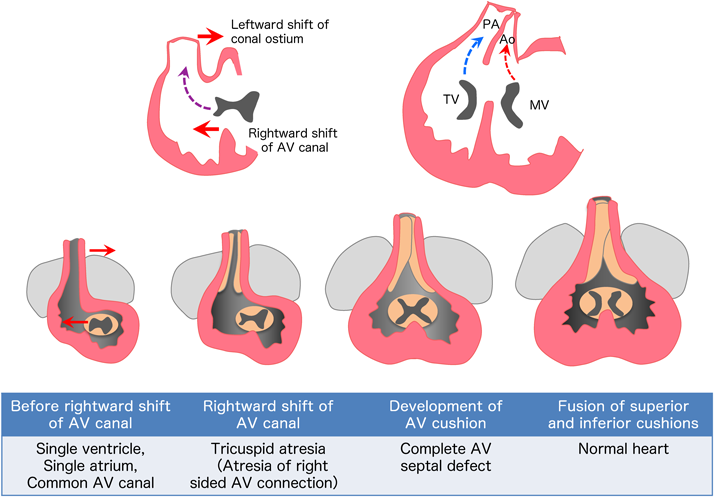
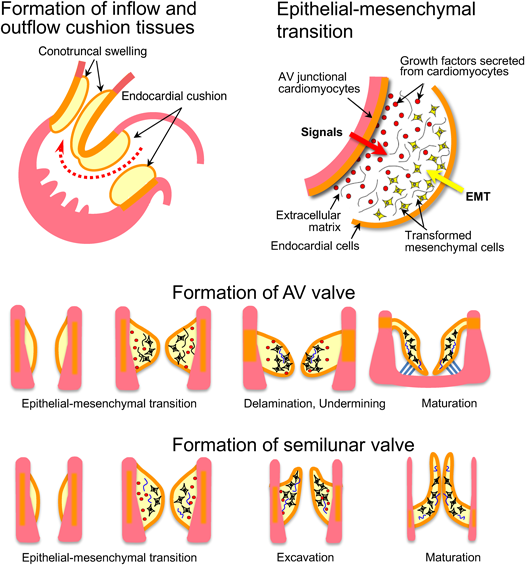
As the looping process progresses, both ventricles bulge and the ventricular septum grows in the middle of the ventricles. As the right ventricle develops, the ventricular septum shifts relatively to the left side of the heart, such that the right side of the atrioventricular connection is newly formed (rightward shift of the atrioventricular canal). After that, the upper and lower endocardial cushions coalesce in the center, dividing the atrioventricular canal into the mitral and tricuspid valves5) (Fig. 5).
Development of Endocardial Cushion Tissue and Formation of Atrioventricular Valve: Common Atrioventricular Canal, Complete and Incomplete Atrioventricular Septal Defect, Ventricular Septal Defect, etc.
In the atrioventricular canal and the conotruncal regions, differentiation of cardiomyocytes is suppressed to form valve tissues. Signaling molecules that promote epithelial-mesenchymal transition (EMT) are secreted from myocardial and endocardial cells. Abundant interstitial tissues develop in the areas of endocardial cushion and conotruncal swelling (Fig. 6). In the inflow tract, four endocardial cushion tissues develop. Mitral and tricuspid valves are formed through a remodeling process called “undermining”. On the other hand, two (initially four) conotruncal swellings develop in the outflow tract. Interestingly, proximal and distal swellings coalesce in a spatially twisted manner, such that the pulmonary artery and aorta are divided in a spiral manner.2–6)
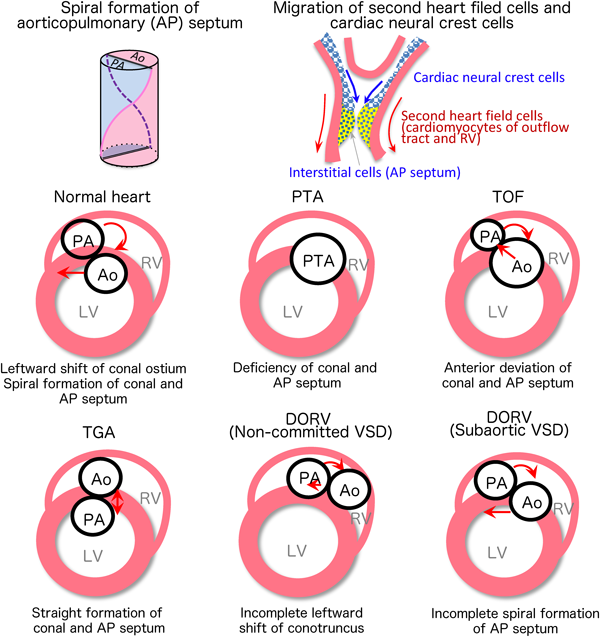
Leftward Shift of the Conal Ostium and Spiral Formation of the Great Arteries: Persistent Truncus Arteriosus, Double Outlet Right Ventricle, Complete Transposition of the Great Arteries, Tetralogy of Fallot, Aorto-Pulmonary Window, etc.
Just after the rightward shift of the atrioventricular canal, the outflow tract and the conotruncus starts to shift to the left side of the heart (leftward shift of conal ostium).The conotruncus is formed in the outflow tract on approximately embryonic day 28. In the inner surface, conotruncal swellings develop and fuse in a twisting manner, dividing the pulmonary artery and aorta into a spiral direction. As a result, the aorta, which is a posteriorly developed great artery, newly connects to the left ventricle. The infundibular septum, which is located at the bottom of the conotruncal swelling, eventually joins to the muscular ventricular septum. Finally, the pulmonary artery and aorta are aligned and connected with the right and left ventricles, respectively. This formation of the right ventricular outflow tract is largely associated with cardiomyocytes (Tbx1, Isl1-positive) derived from the second heart field that migrate and differentiate from the visceral mesoderm. On the other hand, the spiral division of the arterial septum is associated with cardiac neural crest cells (Pax3, Wnt1-positive) that migrate from the cervical neural crest2–5) (Fig. 7).
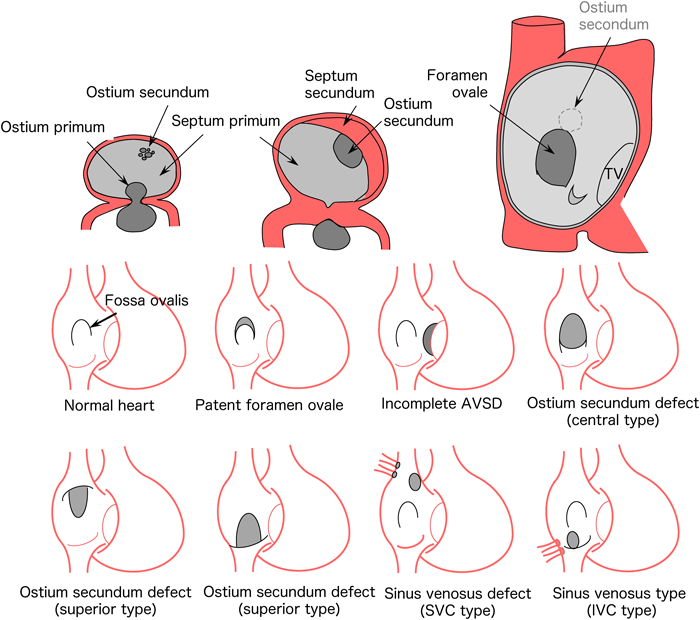
Atrial and Ventricular Septum Formation: Atrioventricular Septal Defect, Atrial Septal Defect, Ventricular Septal Defect
The upper center of the atrium is recessed downward, with the formation of the septum primum on approximately embryonic day 30. It extends downward and leaves a defect hole in the lower end (ostium primum), which is later closed by endocardial cushion tissue. Multiple defects are formed at the higher portion of the septum primum, which assemble to form ostium secundum. As the bilateral atria become bulged, septum secundum is formed on approximately embryonic day 35 to 37. The septum secundum, like the septum primum, leaves a defect hole in the center to the lower end (foramen ovale). The thin septum primum works as a valve between the right and left atria to make unidirectional blood flow from the right to the left atrium. When the baby starts to breathe, a large amount of pulmonary venous blood returns to the left atrium, causing the primary septum to be compressed from the left atrium, which enhances the closure of the foramen ovale (Fig. 8). The classifications of atrial septal defects include patent foramen ovale, ostium primum defect, ostium secundum defect (upper edge defect, foramen ovale defect, lower edge defect), and sinus venosus defects (superior vena cava defect, inferior vena cava defect). Sinus venosus defects are thought to be induced by fusion of the right upper or lower pulmonary veins and superior or inferior caval veins in the early stages of heart morphogenesis.2–5)
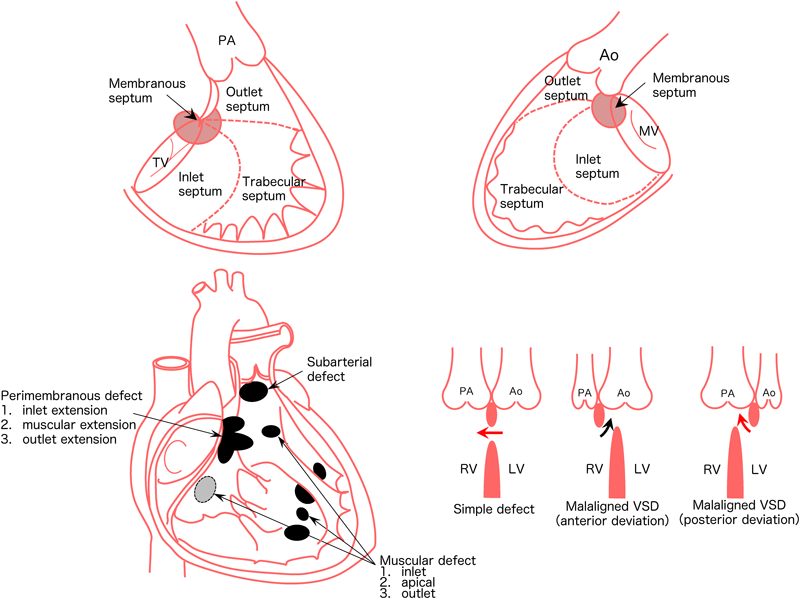
The ventricular septum consists of the following four components. Inlet septum, trabecular septum, infundibular septum, and perimembranous septum. Any abnormal formation of these components induces ventricular septal defects (Fig. 9).
The classifications of ventricular septal defects include:
- Perimembranous defects
- Inlet extension
- Muscular extension
- Outlet extension
- Muscular defects
- Inlet muscular
- Apical muscular
- Outlet muscular
- Subarterial defects
In terms of frequency, perimembranous defects are the most common, accounting for approximately 50% of ventricular septal defects. Septal defects are also classified into two types: simple defects and malaligned defects.1, 5)
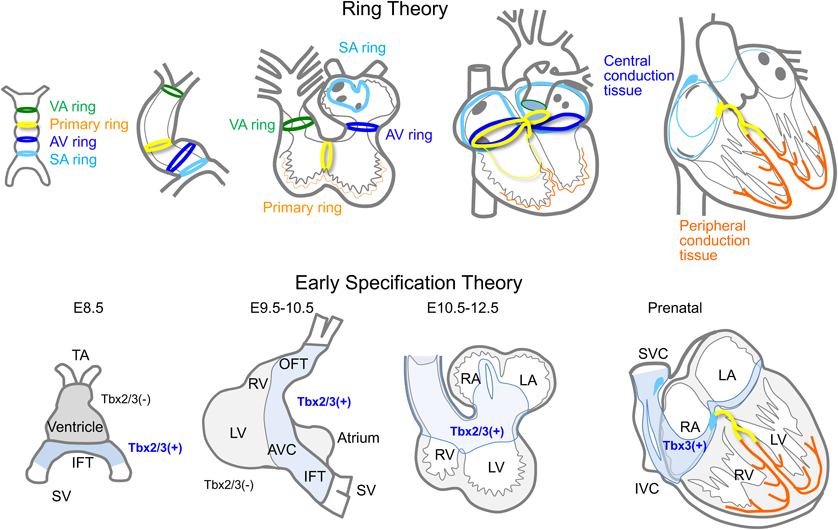
Development of the Cardiac Conduction System
During the development of the cardiac conduction system, various morphogenic or transcription factors are expressed and regressed in a temporally and spatially controlled manner. These factors create a series of conduction tissues from the sinus node to the Purkinje fibers. It has been hypothesized that there are four ring-shaped tissues in the primitive heart tube that eventually give rise to the cardiac conduction system. These ring tissues twisted in a complex manner with looping process and eventually form a central conduction system (Ring theory).7) It appears that Notch and Bmp, along with T-box transcription factors are expressed in specific parts of the atrial and ventricular myocardium from an early stage. They suppress the differentiation of immature cardiomyocytes to mature working myocytes, forming a proximal portion of conduction tissues to the His bundles (early specification theory).8) On the other hand, Purkinje fibers, a peripheral portion of the conduction system, develop with signals from surrounding vascular and interstitial tissues (recruitment). Finally, the proximal and peripheral portions of the conduction tissues connect at the distal end of the His bundles to form a complete one (Fig. 10).
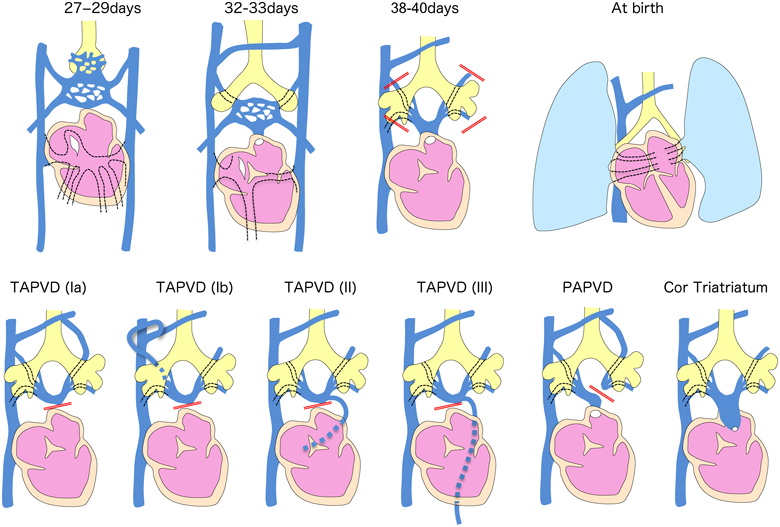
Pulmonary Vein Formation: Total and Partial Anomalous Pulmonary Venous Drainage, Cor Triatriatum, Pulmonary Vein Atresia, etc.
The pulmonary venous plexus develops around the lung bud, which sprouts from the frontal portion of the foregut. The pulmonary venous plexus initially communicates with a systemic venous plexus called splanchnic plexus. On approximately embryonic day 28, the common pulmonary vein protrudes from the posterior wall of the left atrium to form a connection with the pulmonary venous plexus. After that, the communication between the pulmonary venous and splanchnic plexus disappears. If this connection is interrupted, the pulmonary venous blood is unable to drain into the left atrium, resulting in it unavoidably draining into the systemic veins (superior vena cava, coronary sinus, or portal veins). This condition induces total (or partial) abnormal pulmonary venous drainage2–5) (Fig. 11). If the connection between the common pulmonary vein and the left atrium is small, cortriatratum develops. It has recently been clarified that cardiac inflow tract myocardial progenitor cells and pulmonary vascular plexus cells are derived from common progenitor cells (cardiopulmonary mesoderm precursors; CPPs cells), suggesting a relationship with the pathogenesis of pulmonary venous drainage abnormalities.9)
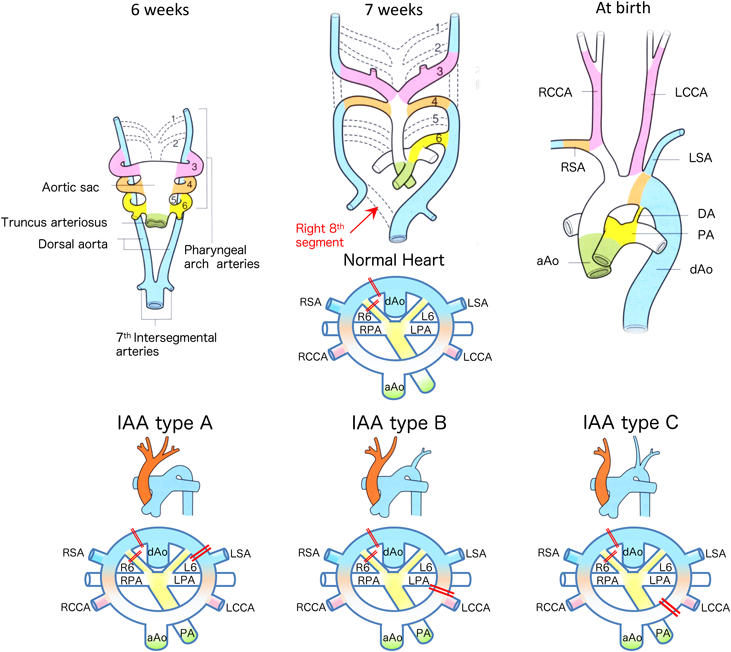
Aortic Arch Remodeling: Interrupted Aortic Arch, Aberrant Subclavian Artery, Vascular Ring, etc.
During the embryonic stages, blood flow from the heart is supplied to the entire body via the common artery trunk, aortic sac, six pairs of pharyngeal arches, and the dorsal aorta. Only the 3rd, 4th, and 6th pharyngeal arches form the aortic arch in humans, with the others disappearing through the remodeling process. The 8th segment of the right dorsal aorta initially regresses and disappears, which initiates remodeling of the human normal left aortic arch. Edward et al. proposed a schematic diagram that explains the development of normal and abnormal aortic arch formation, based on a double aortic arch as the basic form2–5) (Fig. 12). With this schematic diagram, almost all abnormalities of the aortic arch can be clearly explained.
The morphogenesis of the heart is completed in the spatiotemporally-controlled manner shown in this review. The expanding scientific information on embryonic heart development is essential for the diagnosis and treatment of congenital heart disease. Thus, in order to understand the mechanisms of the disease, prevent the initial onset of the disease, and finally improve the quality of life of patients, we must keep investigating the etiology and pathogenesis of congenital heart disease. The information is also essential for the development of regenerative medicine, including the repair and preproduction of cardiovascular tissues.
Conflicts of Interest
The author declares no conflicts of interest in association with the present study.
- CHD
- congenital heart disease
- RA
- right atrium
- RV
- right ventricle
- LA
- left atrium
- LV
- left ventricle
- TV
- tricuspid valve
- MV
- mitral valve
- AV canal
- atrioventricular canal;
- TA
- truncus arteriosus
- SV
- sinus venosus
- IFT
- inflow tract
- OFT
- outflow tract
- RSA
- right subclavian artery
- RCCA
- right common carotid artery
- LCCA
- left common carotid artery
- LSA
- left subclavian artery
- DA
- ductus arteriosus
- aAo
- ascending aorta
- dAo
- descending aorta
- PA
- pulmonary artery
- LPA
- left pulmonary artery
- RPA
- right pulmonary artery
- SA ring
- sinoatrial ring
- AV ring
- atrioventricular ring
- VA ring
- ventriculoarterial ring
- AVC
- atrioventricular canal
- ASD
- atrial septal defect
- VSD
- ventricular septal defect
- AVSD
- atrioventricular septal defect
- SA
- single atrium
- SV
- single ventricle
- AV concordant
- atrioventricular concordant
- TA
- tricuspid atresia
- PA/IVS
- pulmonary atresia with intact ventricular septum
- HLHS
- Hypoplastic left heart syndrome
- TOF
- tetralogy of Fallot
- PTA
- persistent truncus arteriosus
- DORV
- double outlet right ventricle
- TGA
- transposition of the great arteries
- ccTGA
- congenitally corrected transposition of the great arteries
- PAPVD
- partial anomalous pulmonary venous drainage
- TAPVD
- total anomalous pulmonary venous drainage
- IAA
- interrupted aortic arch
- RAA
- right aortic arch
- AS
- Aortic stenosis
- PS
- pulmonary stenosis
- WPW
- Wolf-Parkinson-White
Originally published in Pediatric Cardiology and Cardiac Surgery, Vol. 34 (2018), No. 3, pp. 88–98
引用文献References
1) Sizarov A, Baldwin HS, Srivastava D, et al: Development of the heart: Morphogenesis, growth, and molecular regulation of differentiation, in Allen HD, Shaddy RE, Penny DJ, et al (eds): Moss and Adams’ Heart Disease in Infants, Children, and Adolescents Including the Fetus and Young Adult, 9th edition. Philadelphia, Wolters Kluwer, 2016, pp 1–54
2) Sadler TW: Cardiovascular system. Langman’s Medical Embruyology, 13th edition. Philadelphia, Wolters Kluwer, 2015, pp 175–217
3) Moore KL, Persaud (translated by Seguchi H and Kobayashi T): Cardiovascular system. Moore anthropology. 8th edition. Gracia del Saz E. pp 271–318
4) Dews U (translated by Shiota K): Embryology Atlas. Bunkodo, Tokyo, 1997, pp 132–205
5) Yamagishi H, Shiraishi I: Clinical cardiac embryology to understand congenital heart disease. Medical View Co. Ltd., Tokyo, 2007
6) Shiraishi I, Ichikawa H: Human heterotaxy syndrome: From molecular genetics to clinical features, management, and prognosis. Circ J 2012; 76: 2066–2075
7) Jongbloed MR, Mahtab EA, Blom NA, et al: Development of the cardiac conduction system and the possible relation to predilection sites of arrhythmogenesis. SciWorldJ 2008; 8: 239–269
8) Christoffels VM, Moorman AF: Development of the cardiac conduction system: Why are some regions of the heart more arrhythmogenic than others? Circ Arrhythm Electrophysiol 2009; 2: 195–207
9) Peng T, Tian Y, Boogerd CJ, et al: Coordination of heart and lung co-development by a multipotent cardiopulmonary progenitor. Nature 2013; 500: 589–592
 Isao Shiraishi
Isao Shiraishi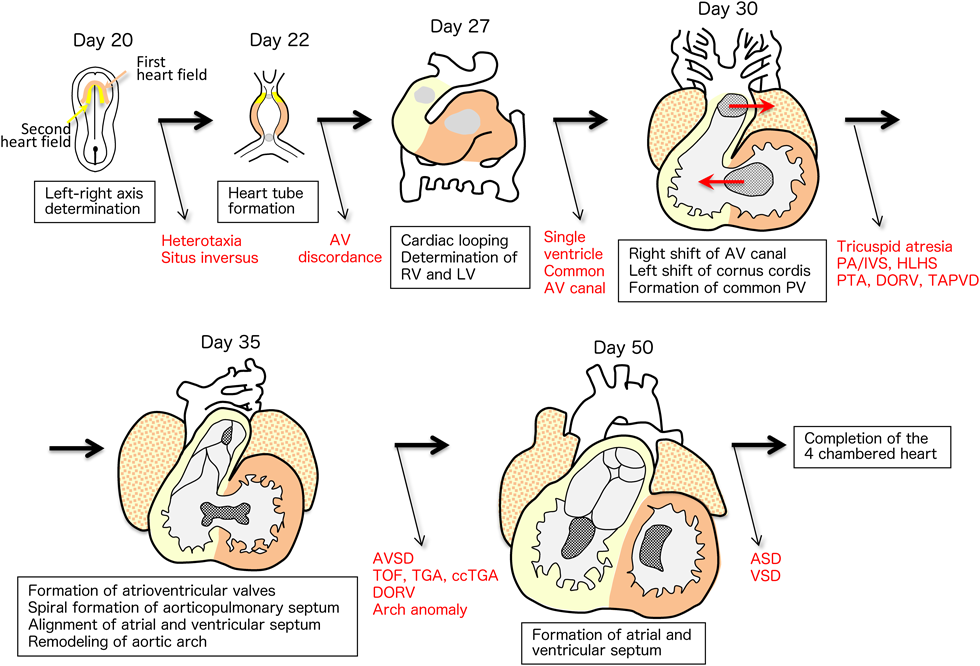
 =first heart field
=first heart field  =second heart field. Left and right atria are derived from the first and the second heart fields.
=second heart field. Left and right atria are derived from the first and the second heart fields.